

| << Chapter < Page | Chapter >> Page > |
No discussion of the evolution of plants on land can be undertaken without a brief review of the timeline of the geological eras. The early era, known as the Paleozoic, is divided into six periods. It starts with the Cambrian period, followed by the Ordovician, Silurian, Devonian, Carboniferous, and Permian. The major event to mark the Ordovician, more than 500 million years ago, was the colonization of land by the ancestors of modern land plants. Fossilized cells, cuticles, and spores of early land plants have been dated as far back as the Ordovician period in the early Paleozoic era. The oldest-known vascular plants have been identified in deposits from the Devonian. One of the richest sources of information is the Rhynie chert, a sedimentary rock deposit found in Rhynie, Scotland ( [link] ), where embedded fossils of some of the earliest vascular plants have been identified.

Paleobotanists distinguish between extinct species, as fossils, and extant species, which are still living. The extinct vascular plants, classified as zosterophylls and trimerophytes, most probably lacked true leaves and roots and formed low vegetation mats similar in size to modern-day mosses, although some trimetophytes could reach one meter in height. The later genus Cooksonia , which flourished during the Silurian, has been extensively studied from well-preserved examples. Imprints of Cooksonia show slender branching stems ending in what appear to be sporangia. From the recovered specimens, it is not possible to establish for certain whether Cooksonia possessed vascular tissues. Fossils indicate that by the end of the Devonian period, ferns, horsetails, and seed plants populated the landscape, giving rising to trees and forests. This luxuriant vegetation helped enrich the atmosphere in oxygen, making it easier for air-breathing animals to colonize dry land. Plants also established early symbiotic relationships with fungi, creating mycorrhizae: a relationship in which the fungal network of filaments increases the efficiency of the plant root system, and the plants provide the fungi with byproducts of photosynthesis.
One of the most exciting recent developments in paleobotany is the use of analytical chemistry and molecular biology to study fossils. Preservation of molecular structures requires an environment free of oxygen, since oxidation and degradation of material through the activity of microorganisms depend on its presence. One example of the use of analytical chemistry and molecular biology is the identification of oleanane, a compound that deters pests. Up to this point, oleanane appeared to be unique to flowering plants; however, it has now been recovered from sediments dating from the Permian, much earlier than the current dates given for the appearance of the first flowering plants. Paleobotanists can also study fossil DNA, which can yield a large amount of information, by analyzing and comparing the DNA sequences of extinct plants with those of living and related organisms. Through this analysis, evolutionary relationships can be built for plant lineages.
Some paleobotanists are skeptical of the conclusions drawn from the analysis of molecular fossils. For example, the chemical materials of interest degrade rapidly when exposed to air during their initial isolation, as well as in further manipulations. There is always a high risk of contaminating the specimens with extraneous material, mostly from microorganisms. Nevertheless, as technology is refined, the analysis of DNA from fossilized plants will provide invaluable information on the evolution of plants and their adaptation to an ever-changing environment.
The green algae and land plants are grouped together into a subphylum called the Streptophytina, and thus are called Streptophytes. In a further division, land plants are classified into two major groups according to the absence or presence of vascular tissue, as detailed in [link] . Plants that lack vascular tissue, which is formed of specialized cells for the transport of water and nutrients, are referred to as non-vascular plants . Liverworts, mosses, and hornworts are seedless, non-vascular plants that likely appeared early in land plant evolution. Vascular plants developed a network of cells that conduct water and solutes. The first vascular plants appeared in the late Ordovician and were probably similar to lycophytes, which include club mosses (not to be confused with the mosses) and the pterophytes (ferns, horsetails, and whisk ferns). Lycophytes and pterophytes are referred to as seedless vascular plants, because they do not produce seeds. The seed plants, or spermatophytes, form the largest group of all existing plants, and hence dominate the landscape. Seed plants include gymnosperms, most notably conifers (Gymnosperms), which produce “naked seeds,” and the most successful of all plants, the flowering plants (Angiosperms). Angiosperms protect their seeds inside chambers at the center of a flower; the walls of the chamber later develop into a fruit.
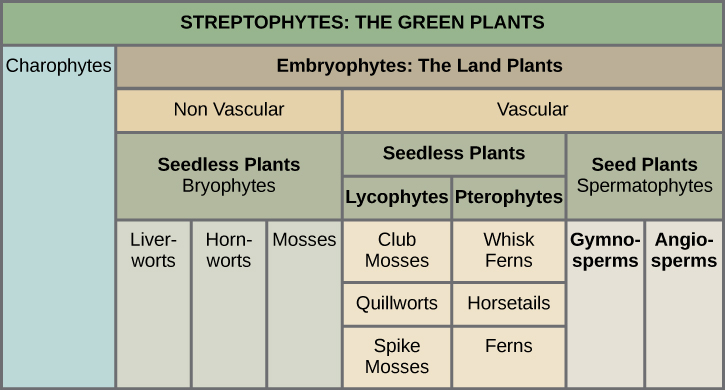
Which of the following statements about plant divisions is false?
Land plants acquired traits that made it possible to colonize land and survive out of the water. All land plants share the following characteristics: alternation of generations, with the haploid plant called a gametophyte, and the diploid plant called a sporophyte; protection of the embryo, formation of haploid spores in a sporangium, formation of gametes in a gametangium, and an apical meristem. Vascular tissues, roots, leaves, cuticle cover, and a tough outer layer that protects the spores contributed to the adaptation of plants to dry land. Land plants appeared about 500 million years ago in the Ordovician period.
[link] Which of the following statements about plant divisions is false?
[link] B.
Notification Switch
Would you like to follow the 'Biology' conversation and receive update notifications?

|
|
|
|
|

|
|

|
|

|
|

|
|

|
|

|
|

|
|

|
|

|
|

|